


Results 1 to 18 of 18
 LinkBack URL
LinkBack URL About LinkBacks
About LinkBacks-
04-30-2002, 09:55 PM #1
 AR-Hall of Famer / Retired
AR-Hall of Famer / Retired

- Join Date
- Aug 2001
- Location
- Wherever necessary
- Posts
- 7,846
 Steroids DO make you agressive!! - Full Study
Steroids DO make you agressive!! - Full Study
32 Copyright © 2001 by MJM
REVIEW ARTICLE
The Role of Testosterone in Aggression
Katherine Simpson * , B.Sc. (Hons.)
ABSTRACT This review article explores the evidence that testosterone is significantly correlated with
certain forms of aggression in a number of animals, although firm evidence is lacking for humans.
Studies have revealed that structures within the limbic system are particularly involved in the
elicitation of aggression and are sexually dimorphic. Testosterone can exert its effects in one of two
ways: either on androgen receptors after conversion to 5-alpha-dihydrotestosterone or on estrogen
receptors after aromatization to estradiol. It can act via genomic mechanisms to induce production of
proteins or via non-genomic mechanisms to modulate neural activity. Androgen and estrogen receptors
are also found along neurotransmitter pathways. As such, testosterone is able to modulate levels of
various neurotransmitters that show evidence of mediating effects on aggressive behaviour. In addition,
recent evidence suggests that these neurotransmitters are involved in processes such as olfaction and
arousal and suggestions have been put forward explaining how testosterone may modulate these
processes. However there is a critical time period early in life, usually within the first few days after
birth, during which testosterone exposure is essential to elicit aggression in adulthood. It is thought that
testosterone and its metabolites sensitize an androgen-responsive system, while estrogenic metabolites
establish the capacity to fight in response to estrogenic stimulation later in life. Despite this, testosterone
is only one of a myriad of factors that influence aggression and the effects of previous experience and
environmental stimuli have at times been found to correlate more strongly.
INTRODUCTION
Changes in the concentration of hormones can have profound effects on mood and behaviour in humans.
Variation in hormone levels have been implicated in some psychiatric disorders such as depression. For
example, 50-75% of patients with major depression show hyperactivity of the hypothalamic-pituitary-adrenal (HPA) axis characterised by excess secretion of cortisol (1,2). Aggressive behaviour in society is a serious social problem. Between 1998-1999, the number of violent offences in England and Wales rose by 6.3%, the majority of which was violence against the person (83%) (3). Although aggression is affected by various influences, there is evidence to suggest that certain biological factors may modulate aggressive behaviour.
This paper reviews the evidence supporting the role of testosterone as a biological factor affecting aggressive behaviour. Human studies have so far demonstrated a relationship between measures of testosterone in adolescent males and aggression (4) with similar results found in women (5). However, results from human studies have also been subjective (6-9) since results are based on psychological questionnaires and observation. For example, girls with a condition called congenital adrenal hyperplasia, in which androgens are secreted in large amounts, have been shown to act in a more rough-and-tumble way during childhood which was thought to correlate with aggression (10). In contrast, strong evidence for the role of testosterone in influencing aggression is available from animal studies.
It is believed that testosterone mediates these effects during two separate periods in life. The first period is the critical time period, usually the first few days after birth during which sensitization of certain neural circuits is thought to occur. The second period occurs in adulthood perhaps through modulation of neurotransmitter pathways and is thought to depend on the earlier sensitization.
This paper briefly reviews the evidence suggesting that testosterone modulates aggression in both males
and females and is followed by a detailed discussion on the evidence which may shed light on the mechanisms involved. As such this paper focuses on effects during the critical time period shortly after birth and also later in adulthood, and also considers the brain regions thought to be involved in the androgen and estrogen responsive pathways. Relevant details regarding neurotransmitter pathways and other factors possibly involved are included to support the evidence.
EVIDENCE FOR A ROLE OF TESTOSTERONE IN AGGRESSION
The term aggression has been defined as a response that delivers noxious stimuli to another person (11). In animals, however, there are various forms of aggression classified into predatory, intermale, fear-induced, irritable, territorial, maternal and instrumental (12). Androgens have been found to affect only certain forms, for example, intermale aggression which can be illustrated by the resident-intruder test. In contrast, predatory aggression is entirely independent of androgens (13). Therefore only the forms of aggression that have been found to have a link with testosterone facilitation will be discussed.
The classical hormone removal and replacement experiment approach has shown a definite link between
testosterone and aggression in animals. In general, castration leads to a decrease in aggression whilst
replacement of testosterone restores the behaviour. However differences in the effects induced by
testosterone have become apparent between males and females. Different regions in the brain modulate
different hormone-dependent aggression (14) and females require longer exposure to androgens after
ovariectomy to induce male-like behaviour (15).
Male aggression
In 1849 Berthold described behavioural changes in cockerels after castration and how these were reversed
after the testes were replaced. Similarly it has been shown that aggression in castrated rats was restored to
its previous levels by androgen replacement (16). More recently, intermale aggressive behaviour in rats induced by electrical stimulation of the hypothalamus was enhanced by androgens (13). Studies in rats, mice, monkeys and humans show that competitive or intermale aggression increases at puberty, whereas it is reduced by castration and increased by testosterone injection (13,16-22).
Female aggression
Aggression in females, like in males, appears to be facilitated by testosterone in a dose-dependent manner
(16). In one study, ovariectomized female rats were given daily injections of testosterone, estradiol or a
placebo. It was found that testosterone increased aggressiveness, measured by the frequency of fighting,
whereas estradiol or a placebo had little effect (23). In general, female aggression patterns were similar to
those seen in males, i.e. ovariectomy without hormone replacement decreased interfemale fighting and
replacement of gonadal testosterone stimulated it (24). However, according to the authors, the presence of
testosterone was not sufficient to activate the aggression. Instead, the missing essential factor was
likely to be an experiential event, for example, competition. Therefore it is possible that hormones are
required for its persistence but are not sufficient to activate the neural circuitry required for aggression.
HOW DOES TESTOSTERONE CAUSE AGGRESSION?
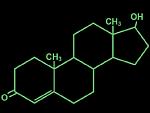
Testosterone acts as a prohormone which when converted into 5-alpha-dihydrotestosterone (5a-DHT)
acts on androgen receptors or when converted into estradiol by the enzyme aromatase, acts on estrogen
receptors (Figure 1). There is overwhelming evidence that most of the effects of testosterone in mediating
aggression occur after aromatization (33). For example, testosterone induced aggression is concurrent with an elevated level of aromatization and nuclear estrogen receptor activity in the hypothalamic/preoptic area.
Treatment with an aromatase inhibitor blocked this aggression and lowered nuclear activated estrogen
receptors (34,35). Furthermore the intensity of aggressive behaviour was directly correlated with the
aromatase activity (p < 0.02) in the posterior hypothalamus (34).
Testosterone also has effects that are manifested early in life. The neonatal organising actions of testosterone are achieved by stimulating cell growth and differentiation in the preoptic area, ventromedial hypothalamus and amygdala, and dendritic branching in the preoptic area and the hypothalamus (35). For example, authors have suggested that testosterone increases the size of the granule cell layer in the dentate gyrus (origin of hippocampal mossy fibres) and that these effects occur during a critical time period (36,37).
Critical time period
The idea that there is a critical time period during which testosterone acts on the brain to sensitize certain
neural circuits has received a great deal of attention and testosterone is now considered to have a definite role in the establishment of aggressive behaviour in adulthood (38-40). Furthermore, it appears that while testosterone modifies aggression, it does so only within a range that has been genetically programmed (41).
In mice, male castration at birth decreased levels of fighting even in the presence of large amounts of
testosterone propionate as adults. However, if the castration took place at ten days of age, the difference in
fighting levels compared to controls was much less pronounced (39). Similar findings were found in
females (16). It was concluded that androgen stimulation of the mouse before ten days of age causes
differentiation of a neural system for aggression which is sensitive to androgens in adulthood. This agrees with others findings (42,43) that castration on day 0 followed shortly by a single injection of testosterone
results in a relatively normal adult male with respect to fighting behaviour.
Recently it has been proposed that in males during the first few days after birth, testosterone and its
metabolites sensitize an androgen-responsive system while estrogenic metabolites organize an estrogen-responsive pathway (44). In general the organizing effect is primarily done by testosterone and its ability to
be aromatized, and can influence either androgen- or estrogen-sensitive circuits (45). For example,
administration of testosterone or methyltrienolone (R1881), a potent nonaromatizable androgen, on day
one of life caused aggression which was brought on in response to androgens but not estrogens in adulthood (46). In contrast, if estrogenic metabolites were present early, then in adulthood this changed with both testosterone or estrogen able to affect aggressive behaviour in males (46).
In females, there is also an androgen-sensitive pathway, although a greater exposure to androgens is
required to induce male-like fighting behaviour. Evidence to support this comes from the finding that
400 mg of testosterone propionate will effectively organise female aggression at 0-2 hours after birth but
it will not at two days of age. However if 600 mg is given at this later stage, the treatment is effective (47).
Furthermore, adult females are completely insensitive to the aggression-promoting effects of estrogens seen
in males.
Therefore there appears to be a sensitive period when the differentiation of aggression with hormones is most likely, i.e. shortly after birth, but the period can be extended by increasing the hormone dosage.
Furthermore the results indicate that hypothalamic and associated brain mechanisms become less sensitive to exogenous androgen with time after castration (39), most likely due to decreased sensitivity of the receptors with age (48). There is, however, some conflicting evidence to suggest that the critical time period is not essential to elicit aggression in adulthood. For example, males castrated on day 0 were found to show aggression in adulthood in response to testosterone however larger doses and often a longer duration of treatment was required (16).
Recently it has been speculated that changes also occur in utero: the testosterone secretory capacity of
Leydig cells in the testes of mice changes differentially during intrauterine development in males of the
aggressive and non-aggressive selection lines (49). The peak secretory capacity is reached at day 17 of gestation for the males of the aggressive selection line while the peak for the non-aggressive line is reached on the first neonatal day. Similarly female mice fought significantly sooner following the initiation of testosterone treatment in adult life if they were exposed to it between the 13 th and 18 th day of fetal development (50). Similar results were obtained by others (51,52). Coincidently, the brain aromatase activity changes during development as well (53-55); for example, at day 1, aggressive males show
higher aromatase activity in the amygdala than in the hypothalamus, whereas in less aggressive strains the
activity did not differ (54).
Figure 1. The chemical structures of testosterone and its derivatives are shown. Testosterone is converted into 5a-DHT by the enzyme 5a-reductase to then act on androgen receptors. Alternatively, aromatase can convert testosterone into estradiol which acts on estrogen receptors.
The effects of testosterone in adulthood
While the primary action of steroid hormones is to regulate protein synthesis through a genomic action,
they can also modulate neural activity via non-genomic mechanisms such as modifying the permeability of
nerve cell membranes. This suggests the existence of membrane receptors for the steroid in addition to
nuclear receptors. These non-genomic effects result in changes in electrical potential possibly through cyclic adenosine monophosphate activation and the release of neurotransmitters into the synapse (56). One of the primary actions of the steroid hormones thus may be to regulate the opening of ion channels in the cell membrane.
It is widely accepted that steroid hormones modulate behaviour through indirect actions on neuro-transmission in the central nervous system. It is assumed that their activity is related to the density of
steroid receptors in these areas (57). When the distribution of steroid hormone receptors in the brain is
compared with the distribution of neurotransmitter pathways, a close relationship is observed (Figure 2);
for example, the distribution pattern of estrogen receptors correlates well with that of noradrenaline,
dopamine and serotonin (5-HT) nerve pathways in the rat (58). When a steroid hormone alters the electrical
activity of a target neuron along one of these pathways the amount of neurotransmitter released from that
neuron is altered. For example, it was found that normal fighting in mice decreased when there was an
accelerated release of certain amines (59).
However it should be stressed that the neural circuits underlying aggression are themselves plastic, i.e.
neurotransmitter modulation may not be the only way testosterone can affect aggression in adulthood. For
instance, levels of aggression tend to fluctuate during the year in many animals according to their mating
season: when the breeding season approaches and the gonads recrude, aggression increases with a
corresponding elevation in testosterone levels (60).
Effects of testosterone on neurotransmitter pathways
Many studies have been undertaken that implicate 5-HT in modulating aggression. In general, increased
activity of serotonergic synapses inhibits aggression: studies in male rhesus monkeys showed that those with low levels of 5-hydroxyindoleacetic acid (5-HIAA) were more aggressive (30,61,62). Furthermore,
fluoxetine, a serotonin reuptake inhibitor, tends to decrease aggression in both animals and humans. Strong
evidence suggests that these effects are mainly mediated via 5-HT 1A and 5-HT 1B receptors (63). This is
supported by various animal studies: 5-HT 1A agonists decrease aggression and defensive behaviour in wild rats (64); gene knockout mice lacking the 5-HT 1B receptor (equivalent to 5-HT 1Db in humans) showed greatly enhanced intruder confrontation aggression (65).
Testosterone may also be involved in the arginine vasopressin (AVP) system, since an AVP receptor
antagonist injected into the anterior hypothalamus of male golden hamsters was found to block their
offensive aggression (66). This is supported by evidence that male golden hamsters have androgen-sensitive AVP receptors in the ventrolateral hypothalamus and castration leads to a loss of AVP
binding (67). Furthermore, there appear to be 5-HT synapses on AVP neurons in the anterior hypothalamus
and the AVP immunopositive boutons and hippocampo-septal axon terminals frequently terminate on the same somatospiny neurons on the lateral septum. Therefore, perhaps testosterone promotes aggression by lowering 5-HT levels thus removing a potential inhibitory influence on AVP.
Testosterone has also been found to influence the density of GABA A receptors. Testosterone-treated rats
have a decreased density of GABA A receptors in several brain regions (68) and several studies have
implicated low levels of GABA with aggression (69,70). Coincidently, a role for GABAergic inhibition
has been implicated in odour coding and in particular GABA A antagonists caused lowering of the response
threshold to odours (70,71). Therefore high-attacking animals would have a better perception of the olfactory cues leading to rapid recognition of the opponent. There is also evidence to support a direct role of noradrenaline in aggression (72). Adrenoceptor blocking drugs such as phentolamine (alpha receptor
blocker) or obsidan (beta blocker) caused a decrease in aggression and testosterone levels in dominant mice
(73). It has been suggested that noradrenaline heteroceptors terminate on presynaptic 5-HT neurons
raising the possibility of interaction between the two transmitter systems (72).
Finally, testosterone has been found to suppress dopamine turnover in the anterior hypothalamus of
male rats (74) which was thought to be involved in androgen-dependent aggression, although studies tend
to support a positive relationship between central dopamine function and aggression (72). There have also
been suggestions that acetylcholine and glutamate are involved in testosterone-mediated aggression.
OTHER FACTORS INVOLVED IN TESTOSTERONE-MEDIATED AGGRESSION
Research into human forms of behaviour such as aggression is complicated by the complex manner in
which humans tend to behave. For example, studies have found that aggressive encounters are influenced by learning, i.e. the outcome of the first encounter influenced subsequent fights (75). There is also
evidence that aggression is in itself rewarding. For example, when animals with lateral hypothalamic
electrodes were allowed to control the stimulation, they pressed the levers with high frequencies suggesting the presence of positive feelings (76). Interestingly, self- stimulation rates were decreased when the animals were castrated and increased again after hormone substitution.
Other research has indicated that testosterone may be correlated with aggressive motives and competitiveness rather than violence per se. Serotonin function on the other hand may function to limit aggression to an appropriate time, setting and intensity. Overall the results indicate that while subjects with high levels of testosterone exhibit more aggression, the aggressive encounters are stopped before they escalate into dangerous forms of aggression (77). Other factors implicated in aggression include involvement of the Y chromosome (78,79), alcohol (80) and the role of olfaction (81-84).
Figure 2. Neural circuits underlying aggression as shown in the rat brain. Original work by Hess in 1928 (26) found that sham rage occurred after electrical stimulation of the hypothalamus. More recently, a continuous stretch of points through the hypothalamus have been mapped out in rats which when stimulated resulted in aggressive behaviour (27). Furthermore it appeared that for each form of aggression, there appeared to be different representations within the hypothalamus. For example, stimulation and lesion studies point to a strong lateral hypothalamic involvement in predatory and intermale aggression whereas the anterior hypothalamus is involved in fear-induced and irritable aggression. As such the neural systems underlying these different forms of aggression seem to converge from various parts of the brain within the hypothalamus (28). Various lesion, electrophysiological and morphological studies suggest that the neural control of aggressive behaviour, in particular the associated movements, are programmed by neural circuits in the brain stem whose activity appears to be controlled by various structures in the limbic system such as the hypothalamus and amygdala (29,30). In turn, important pathways lead from the hypothalamus through the medial forebrain bundle to the limbic cortex through the amygdala and septum. There are also connections from the olfactory bulb (31). Interestingly, high densities of androgen and estrogen receptors were found in these areas (32). Schematic diagrams show parasagittal views of the rat brain at a medial (A) and lateral (B) position. The neural circuits thought to be involved in aggressive behaviour are indicated as dotted lines. Dots represent areas found to contain androgen and estrogen receptors. Notice the overlap between the indicated neural circuits and the regions of highest androgen and estrogen receptor density.
PROBLEMS WITH PREVIOUS STUDIES
Most studies of testosterone and aggression have measured plasma total testosterone levels which can be
problematic since in plasma the free fraction accessible to the central nervous system is only about 2% (85). It was therefore proposed that accurate reflection of central androgen activity might be better obtained by cerebrospinal fluid (CSF) free testosterone (77). Furthermore, investigators found that in rhesus
monkeys testosterone was related to overall aggressiveness whereas low levels of CSF 5-HIAA
were related to impulsivity. This has implications on a variety of studies where impulsivity was regarded as a predisposition or even a type of aggression.
In addition, there are species differences in the structures involved in the different types of aggression.
The cingulate gyrus is involved in fear-induced aggression in the monkey but irritable aggression in the
cat and dog. The effects of stress are also problematic since they have been found to reduce testosterone in
man and other animals. Therefore stress induced by defeat would naturally depress testosterone levels (86).
Related to this, it was found that sexual stimulation (87) and exercise (88) increase testosterone levels and many of the behaviours elicited in the studies probably have an element of sexual motivation which has been shown to be estrogen dependent (89).
Although there is strong evidence that testosterone is involved in aggression, it must be stressed that perhaps other hormones are involved. After castration, many hormone systems are disrupted which could confound the experiment. For example, aggression was more closely related to luteinizing hormone than testosterone in the male starling (90). Review and clarification of behavioural measures is therefore recommended.
CONCLUSION
There is strong evidence in animals that testosterone is directly associated with aggression, although this
correlation is not as strong in humans. Testosterone appears to mediate its effects during a critical time
period shortly after birth during which it sensitizes certain neuronal circuits in the brain. As a result, in
adulthood, when these circuits are stimulated again by steroids , aggressive behaviour is elicited perhaps
through modulation of specific neurotransmitter pathways. However, it must be remembered that
hormones themselves do not directly cause behaviours, but induce chemical changes in certain neurons,
affecting the likelihood of certain behavioural outcomes as a result of modulation of particular neural pathways (91). Furthermore, gonadal hormones are only one of a myriad of influences on aggressive behaviour. Since testosterone is present in males that are not aggressive as well as in those that are, it is obvious that another factor(s) is involved, such as cognition and environmental circumstances which have been found to affect the expression of aggression (60,92).
Scientists are limited in that they cannot perform experimental procedures in humans as they have done
in animals and since the behaviour patterns in humans are so different from animals, rendering it difficult to
extrapolate from animals to humans. As it was put recently there is a danger of triviality and truism, or
even misleading simplification, in many of our extrapolations and animal models (76). As such we
should remember that correlation does not necessarily indicate causation and in many studies such
assumptions have been made. Before experiments can be done to investigate these correlations, we need to be sure that the types of aggression we are looking into are testosterone-dependent and eliminate other factors that could confound the results.
-
04-30-2002, 10:12 PM #2
 Banned
Banned
- Join Date
- Jan 2002
- Location
- Miami, Fla
- Posts
- 4,744
Sticky material, bump
-
02-13-2005, 11:15 PM #3
 Member
Member
- Join Date
- Feb 2005
- Location
- crashing my boat
- Posts
- 742
long
-
02-13-2005, 11:18 PM #4
 Member
Member
- Join Date
- Oct 2004
- Location
- Arkansas
- Posts
- 854
you'll never learn anything if you dont read the long studies, those are the juicy ones =) Originally Posted by rager
Originally Posted by rager
Alex
-
02-13-2005, 11:19 PM #5
 Senior Member
Senior Member
- Join Date
- Mar 2004
- Location
- Injecting in the bathroom
- Posts
- 1,360
This **** is 3 years old..
What the hell are you doing?
-
02-13-2005, 11:20 PM #6
 Senior Member
Senior Member
- Join Date
- Oct 2004
- Posts
- 1,216
Good info. Bump this to the top!
-
02-14-2005, 12:38 AM #7
 Senior Member
Senior Member
- Join Date
- Feb 2003
- Location
- CeN CAL
- Posts
- 1,222
I think if u have taken juice you would no this allready!!!
-
02-14-2005, 12:44 AM #8
 Anabolic Member
Anabolic Member
- Join Date
- Feb 2004
- Posts
- 3,651
Originally Posted by rager
Way to bump a nearly 3 year old thread and then bitch about it Bro")
-
02-14-2005, 04:52 AM #9
 Senior Member
Senior Member
- Join Date
- Dec 2003
- Location
- South JerseyJ
- Posts
- 1,223
Thanx for the bump Great read
-
02-14-2005, 06:25 AM #10
 Member
Member
- Join Date
- Dec 2004
- Location
- UK
- Posts
- 871
Bumpity Bump...
-
02-14-2005, 08:35 AM #11
Senior Member
- Join Date
- Apr 2004
- Posts
- 1,381
Great read, but all studies done in animals.
Also, being a 4 year old study, they now probably think steroids have a high likelihood of shooting bananas out your ass or something totally different like that.
-
02-14-2005, 09:42 AM #12
 Associate Member
Associate Member
- Join Date
- Jan 2004
- Posts
- 432
true Originally Posted by needle
-
02-15-2005, 01:33 AM #13
 Elite Hall Of Fame ~ RIP ~
Elite Hall Of Fame ~ RIP ~
- Join Date
- Dec 2001
- Location
- In a cage near you...
- Posts
- 4,721
Originally Posted by powerlifter

-
02-15-2005, 12:07 PM #14
 Associate Member
Associate Member
- Join Date
- Dec 2004
- Posts
- 249
bump
-
02-16-2005, 01:00 AM #15
Member
- Join Date
- Feb 2005
- Location
- crashing my boat
- Posts
- 742
I did read... nice .. just long Originally Posted by ArkansasAlex
-
02-16-2005, 01:02 AM #16
 Junior Member
Junior Member
- Join Date
- Jan 2005
- Posts
- 138
Good post. It was a lot to read though.BUMP
-
02-16-2005, 01:08 AM #17
Member
- Join Date
- Feb 2005
- Location
- crashing my boat
- Posts
- 742
Originally Posted by needle
lol no kidding? who doesnt know this S?
-
02-16-2005, 01:09 AM #18
Member
- Join Date
- Feb 2005
- Location
- crashing my boat
- Posts
- 742
what the f.. I didnt post this bro? Originally Posted by powerlifter
 Reply With Quote
Reply With QuoteThread Information
Users Browsing this Thread
There are currently 1 users browsing this thread. (0 members and 1 guests)






cutting/ fat loss advice needed...
04-16-2024, 01:34 AM in ANABOLIC STEROIDS - QUESTIONS & ANSWERS